
No joke: someone stepped in it
 April Fool’s Day is just around the corner. And while this paper in the august journal Annales de Paléontologie isn’t supposed to be a joke, it definitely should be:
April Fool’s Day is just around the corner. And while this paper in the august journal Annales de Paléontologie isn’t supposed to be a joke, it definitely should be:
Miguel Telles Antunesa, Ausenda C. Balbino and Léonard Ginsburg (2006). “Miocene Mammalian footprints in coprolites from Lisbon, Portugal.” Annales de Paléontologie, 92(1), pp. 13-30. January-March 2006.
Abstract: For the first time, at least for the Lisbon Miocene series, uncommon ichnologic evidence has been recognized, i.e. mammalian footprints in coprolites. Three coprolites were recorded in three successive stratigraphic units, IVb and Va2 from the Lower Miocene to Vb from the early Middle Miocene. The largest, tridactyl footprint can be ascribed to a right foot of a rhinoceros. Size excludes all the rhinocerotids known from the Vb unit except Hispanotherium matritensis. A smaller coprolite (Va2 unit) shows a tridactyl, left foot impression of a perissodactyl. It is clearly too small for a rhinoceros, even for a young one. It seems to have been made by an Anchitherium Equid. The pes had a plantar pad as still found in the Mesohippus–Anchitherium lineage but not in more advanced Equids. Both tridactyl imprints may have been produced by the coprolite-makers. A large coprolite (IVb unit) that may have been produced by Brachyodus onoideus shows a few didactyl imprints. An artiodactyl trampled the dung with hoofs sliding on its surface and producing two incomplete imprints. It also trampled the dung in a more stable position, producing the best imprint, whose structure indicates it was produced by the left manus. The lack of lateral toe marks excludes suids (and Brachyodus, also because its size is too much small). It is from a small-sized ruminant, most probably a cervid, genus Procervulus. In all cases, defecation occurred on dry land, albeit in eventually or seasonally flooded areas near a river.
Hat-tip: alert paleontologist Alan Gishlick. For some quotes on the significance and methods of this research, see below the fold.
Introduction
The commercial exploitation of sandpits in or close to Lisbon was a highly important activity from an economic viewpoint. It further contributed to Paleontology as many vertebrate fossils were collected, most often by workers, from late lower and early middle Miocene sand units that correspond to regressive, mainly fluviatile events in an otherwise essentially marine series.
[…]
Sandpit exploitation in the study area was often carried out in very dangerous conditions and many workers perished in accidents. As a consequence, all sandpit exploitation was entirely forbidden in 1967.
Fossil collecting has nearly entirely ceased since then, except for systematic washing and sieving to obtain small mammals as well as fish remnants. Even so, an impressive amount of data about medium- and large-sized vertebrates had already been obtained.
[…]
As commercial exploitation progressed quite fast, no erosion or sedimentary surfaces have been exposed, or at least not long enough to allow footprint prospecting.
We had to try ecologic interpretation on the basis of data concerning the taxa and their relative numbers, as well as sedimentary data. In the lower Tagus basin several transgression and regression events have been recorded. Some of the latter correspond to the deposition of fluviatile sands rich in mammals and other fossils. Coprolites were sometimes found in great numbers. However coprolites were often despised and left aside. Only a few had been collected as samples.
In the later times of sandpit exploitation, one of us (M.T.A.) once found a somewhat broken but especially interesting specimen that contained a rather large fish vertebra. The fish was certainly a Lates, a close relative of the common Nile perch that was then common in association with tropical catfishes, i.e. Arius and Chrysichthys (Antunes, 1989; Antunes and Gaudant, 2003). As the vertebra did not undergo a chewing process and resisted all the way through the digestive tract, the predator had not been a mammal. It could therefore be ascribed to a fish-eater as the then common, large-sized, long-snouted crocodilians Tomistoma lusitanica and Gavialis sp. (Antunes, 1987 and Antunes, 1994).
As a consequence we paid much more attention to coprolites. This policy was rewarded. Several types and sizes are represented, including the specimens under study. All specimens belong to the M. Telles Antunes Collection and are provisionally housed at the Centro de Estudos Geológicos, New University of Lisbon.
The majority of coprolites were produced by mammals. Some can be attributed to large beasts such as rhinoceros; other ones, similar in shape to elephant’s, may be ascribed to mastodons (Deinotherium being too rare, it is most improbable as a coprolite-maker).
Some coprolites may have been produced by Amphicyon or other large Carnivora, smaller ones may be droppings from small artiodactyls. It is not uncommon to find round holes dug by dung-seeking Coleoptera.
No other specimens contained recognizable organic remains, although some displayed what could be interpreted as plant impressions.
Indeed a few coprolites revealed what we thought could be the first recognized trampling by some mammal (not necessarily by their makers).
Let us recall that when a mammal defecates it has his hind limbs and especially his feet close by the faeces it expels. Therefore, it is possible that it could eventually trample on its own excrement with the hind feet rather than with its fore feet. Faeces could otherwise be trampled not by their makers but by another animal, whether from the same species or not.
Coprolite makers are less likely to be very young, suckling animals because faeces are produced in smaller quantities, and tend to be less compact, very fragile, and even quite liquid.
The following text concerns the description and interpretation made as we tried to infer the identity of the coprolite and imprint makers. A further interpretation difficulty lies in the distortions that undergo the faeces subject to trampling; there is evidence indeed of sliding of the feet over the faeces’ surface. Anyway, in all the specimens dealt with here, faeces were still soft, but rather advanced in the fluid loss process either through desiccation or compression. Desiccation is obvious in one specimen with a rather small tridactyl imprint since it presents some thin cracks or fissures.
There are several difficulties to overcome, as trackways and footprints are uncommon and so little-documented in detail. As noted by Demathieu et al. (1984: p. 174), a few Miocene cracks and footprints have been described in Hungary, Romania, Switzerland and Austria. The same orders are represented: Perissodactyla (Rhinocerotoidea), Artiodactyla (by far the commonest) and a few Carnivora (idem, p. 175).
However, all the previously studied sites have footprints in sediments, not coprolites! Hence they are not entirely comparable with our specimens. As far as we can ascertain, it seems we really are dealing with a unique case.
It’s nice to see scientists exhibit such obvious expertise and joy in the subject they are studying.
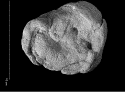
Here is a drawing that reconstructs the difficult-to-interpret photograph at the top of the post:
Fig. 2. Drawing of the coprolite in the same position. The arrow approximately indicates the direction and extension of the foot displacement by sliding on the dung.
PS: I know this is funny, but in the comments, please make artful use of @#*$!%(## rather than just typing the obvious words. We would prefer that PT not be banned by nanny programs and the like.