
Making Babies - How Simple Rules Build Complex Bodies
The New York Academy of Sciences provides us with access to a talk by Nobel Laureate Christiane Nüsslein-Volhard on how genes drive development, no need for unspecified ‘Intelligent Designers’, no need for miracles, just hard work by scientists who are committed to discovering the details of how, what, when and so on. Compare this with how ID explains the development of the embryo.
Click on the Flash presentation
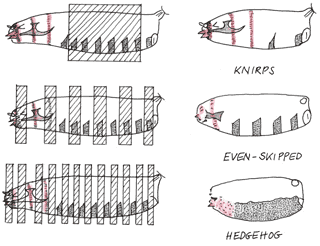
I also suggest that interested readers get their hands on her book “Coming to Life: How Genes Drive Development” by Christiane Nüsslein-Volhard or read an excerpt of the book: Chapter IX — Evolution, Body Plans, and Genomes