
ERV: A Day In The Life of a DI Fellow: Behe
Our friend Abigail Smith aka ERV provides us with an entertaining posting about who else but Behe
Remember Behe’s crusade for Intelligent Design at KKMS 980AM, a Christian radio station?
For those who cannot stand the torture of listening to Michael Behe making claims that science can reliably detect ‘design’, let’s first explain what exactly Intelligent Design is all about and reject ID’s hollow claims.
First of all, how does Intelligent Design define ‘design’? Contrary to what most people may believe, the definition of ‘design’ as used by the ID movement is quite limited and is nothing more than the set theoretic complement of the disjunction of chance and regularity, or in simpler words that which remains when we have exhausted our understanding of scientific explanations. And while science correctly accepts our ignorance, Intelligent Design has chosen to refer to this state of ignorance as ‘design’.
Now things get worse since ‘design’ does not even require an ‘intelligent designer’. What? … Yes, you have heard correctly, and although few ID proponents seem to be aware of this ‘minor detail’ or quickly ‘step over it’, William Dembski made quite a concession when he stated
As reported by Ryan Nichols in his excellent paper “ Scientific content, testability, and the vacuity of Intelligent Design theory “
Before I proceed, however, I note that Dembski makes an important concession to his critics. He refuses to make the second assumption noted above. When the EF implies that certain systems are intelligently designed, Dembski does not think it follows that there is some intelligent designer or other. He says that, "**even though in practice inferring design is the first step in identifying an intelligent agent, taken by itself design does not require that such an agent be posited. The notion of design that emerges from the design inference must not be confused with intelligent agency**" (TDI, 227, my emphasis).
So in other words, Intelligent Design is nothing more than the age old ‘God of the Gaps’ in which our ignorance is somehow seen as evidence for design. In case of ID however, the concept of design is far more constrained and need not even point to intelligent agency.
Wesley Elsberry pointed out this major shortcoming of Intelligent Design’s argument in an early review of Dembski’s work. First of all Wesley recognizes how the term ‘design’ has little relationship to the term as used by earlier Intelligent Design proponents like Paley.
One may wonder what TDI was supposed to accomplish, if design no longer means what Paley meant by it and the attribution of agency no longer follows from finding design. But Dembski believes that finding design does imply agency, even though he has identified that implication as being unnecessary. In his view, because we can often find that design is found where an intelligent agent has acted, we can reliably infer that when we find design, we have also found evidence of the action of an intelligent agent. Section 2.4 gives Dembski’s take on how we go from design to agency. Dembski invokes his explanatory filter as a critical piece of this justification.
Dembski believes that not only design but also agency is found by his argument. This is the message being spread by various and sundry of the “intelligent design” proponents and by Dembski himself in other writings. But is it a secure inference? In his First Things article, and to a lesser extent in his section 2.3 of TDI, Dembski takes biologists to task for avoiding the conclusion of design for biological phenomena. Dembski says that to avoid a design conclusion, biologists uniformly reject one or more of the premises of his argument. But Dembski does not exclude natural selection as a possible cause for events which can be classified as being due to design.
The apparent, but unstated, logic behind the move from design to agency can be given as follows:
- There exists an attribute in common of some subset of objects known to be designed by an intelligent agent.
- This attribute is never found in objects known not to be designed by an intelligent agent.
- The attribute encapsulates the property of directed contingency or choice.
- For all objects, if this attribute is found in an object, then we may conclude that the object was designed by an intelligent agent.
This is an inductive argument. Notice that by the second step, one must eliminate from consideration precisely those biological phenomena which Dembski wishes to categorize. In order to conclude intelligent agency for biological examples, the possibility that intelligent agency is not operative is excluded a priori. One large problem is that directed contingency or choice is not solely an attribute of events due to the intervention of an intelligent agent. The “actualization-exclusion-specification” triad mentioned above also fits natural selection rather precisely. One might thus conclude that Dembski’s argument establishes that natural selection can be recognized as an intelligent agent.
In other words, whenever an ID proponent claims that ‘design has been inferred’, ID cannot even have ruled out scientific explanations like natural selection as the ‘designer’. In other words, it seems that ‘design’ is nothing more than a position of ‘ignorance’ and unless the ID proponents propose their own detailed explanation, ID remains scientifically vacuous.
In other words, we have so far established that the concept of ‘design’ as proposed by ID has little relevance to how the term is more commonly used and interpreted, and that it is a placeholder for our ignorance. We also have established that in order to establish who or what is the ‘designer’, we need to provide detailed explanations. So where are these detailed explanations from Intelligent Design, you may wonder?
Don’t hold your breath, ID is not in the business of responding to such pathetic requirements
William Dembski was asked a very similar question and ‘responded’ that
As for your example, I’m not going to take the bait. You’re asking me to play a game: “Provide as much detail in terms of possible causal mechanisms for your ID position as I do for my Darwinian position.” ID is not a mechanistic theory, and it’s not ID’s task to match your pathetic level of detail in telling mechanistic stories. If ID is correct and an intelligence is responsible and indispensable for certain structures, then it makes no sense to try to ape your method of connecting the dots. True, there may be dots to be connected. But there may also be fundamental discontinuities, and with IC systems that is what ID is discovering.
William A. Dembski Organisms using GAs vs. Organisms being built by GAs thread at ISCID 18. September 2002
All that ID has to offer are ‘fundamental discontinuities’. So what are these so called fundamental discontinuities? Complex Specified Information (CSI) ? Irreducible Complexity? Nope, neither one is able to show that there are fundamental discontinuities that cannot be explained by science. So what else is there? Behe, realizing the vacuity of Intelligent Design so far, has re-attempted to define an ‘edge to evolution’ by taking a guestimate and turn it into a strawman against evolutionary theory. And not even evolutionary theory but just a Darwinian component of such a theory.
However, as shown by scientists like ERV, Behe’s comments lack foundation in reality and are largely based on ignorance. This is where we return to present day. Behe, annoyed that a mere graduate student has shown the vacuity in his position, complained about ERV’s tenacity on the Christian show. Now, this show did not invite ERV for her position but did allow Behe to respond to ERV’s observation that she had gotten Behe to admit that the basic premise of his book had been falsified.
Around 12:00 minutes we hear
Interviewer: “Hey Jeff, I have got Behe to admit that his basic premise to his book “The Edge of Evolution” is wrong. He stated over and over and over that no new protein to protein interactions have ever evolved, ever. And I showed him that one evolved in HIV within the last few decades. And she goes on to say “in his case he is most certainly not ignorant as he admitted defeat. The only logical explanation as to why he is continuing down this current path is that he is a liar”.
What is she talking about and how do you respond
Behe: “Oh clever, eh well uhm she eh she eh is wrong um I um um am not quite eh sure where she is getting this but eh I didn’t say , as matter of fact, I didn’t say that protein sites could not uh evolve, as a matter of fact, I did in my book and that is the sickle cell eh hemoglobin binding site. Now I don’t know if many of your listeners know but [throat clearing sound] in [throat clearing sound] the disease sickle cell anemia eh the hemoglobin which lives in our red blood cells uh kind of gunks things up and uh turns thing into kind of a semi solid mass because the proteins, the hemoglobins stick to eachother. And that is kind of the result of a mutation which is actually useful if you live in malaria affected countries and I said that such things could happen and I said that eh the eh limit for Darwinian evolution was two connected protein-protein binding sites that is if you need three proteins to do something eh and uh each pair of proteins has to be connected by a site you need two protein binding sites, that would be something that based on the data I talked about in the book ‘the edge of evolution’ that is the most you could expect out of eh out of eh eh Darwinian processes. Now, this lady, eh this graduate student said well hey look we have, there’s been a new binding site eh which has occurred in one protein in HIV, the virus that causes aids, hm hm. And she was right and I had not seen that so I said ok well, there is another single binding site that has come about but uh again you would expect that to happen because the odds against a single binding site popping up are not that they are not that prohibitive. But if you need two binding sites together to do something then that’s what I said in my book was pro prohibited. And uh, so since most molecular machines in the cell consist of uh an aggregate of a dozen proteins or so, each of which has to bind very specifically to eachother then it that means that most of the protein machinery in the cell is well beyond Darwinian evolution. So uh so uh she uh, she misread the book uh and she’s uh uh she’s trying to uh she is trying well uh she misread the book. okay ..
Interviewer: Well that is why we need to get the facts
Behe: that’s right that’s right so it uh the the long the short is that most people don’t understand that Aids viruses HIV viruses rather uh occur in enormous numbers like you know billions and billions and billions as Carl Sagan might say.
Interviewer: hm hm
Behe: And they mutate at an enormous rate, tens of thousands of times faster than the cells in our body and and ma mammalian bodies and so on. And yet, after 50 years, uh at this enormous rate uh which I show in my book is uh they have undergone about the same number of mutations as cellular organisms would undergo in billions of years. Uh you have very very little to show for very very little that is new in the HIV virus uh so in my book I try to find the line between what random Darwinian processes can do like Darwinian evolution and what needs design and so certainly there are some little things that that chance can accomplish in in things like HIV , things like malaria which I talk about in the book and and other uh things called ecoli bacteria which I talk about in the book. Some things can be done but like I say there’s there’s quite uh uh strict limit. to to what can happen.
Now remember how ERV explained how Behe had claimed that
“Like malaria, HIV is a microbe that occurs in astronomical numbers. What’s more, its mutation rate is 10,000 times greater than that of most other organisms. So in just the past few decades HIV has actually undergone more of certain kinds of mutations than all cells have endured since the beginning of the world. Yet all those mutations, while medically important, have changed the functioning virus very little. It still has the same number of genes that work in the same way. There is no new molecular machinery. If we see that Darwin’s mechanism can only do so little even when given its best opportunities, we can decisively conclude that random mutation did not build the machinery of life.”
As far as I can tell from Behe’s ‘response’ he agrees with ERV that he was wrong in his book however, he claims that well, a single such binding site might still be probable but two of these are outside the realm of Darwinian evolution.
Read the whole story here where Ian Musgrave hold Behe to the task at hand
It is good to see that you agree that the Golgi targeting sequence is an example of a binding site. However, you don’t get to ignore it because “viral proteins are special”. As I showed in the post you are supposed to be replying to, this is nonsense. In your book you categorically state HIV has developed no new binding sites, the diagram on page 145 of “Edge of Evolution” has a big zero on it. Yet your own example of a binding site is the haemoglobin S mutation, a single amino acid mutation that just clumps up proteins. You don’t write in your book “HIV has evolved several binding sites, but they don’t count because they are viral-protein-host protein interactions” or “HIV has evolved several binding sites, but they don’t count because they are equivalent to the HbS mutation”, you just write zero
However Ian does not end it here and shows how science does explain how multiple binding sites evolve
Now, if you only looked at AII and the related hormone AIII, you would believe that three binding sites RYF, were required for binding. You might puzzle over how you could simultaneously get three replacements in an ancestral, non-binding peptide so that it would bind to this site.
But as we look at SarIle and SarAsp, we can see a way. These have only two of the three binding amino acids present in AII and AIII, yet they bind very well indeed. Maybe only binding to the the DN part of the DNKH motif in the AT-1R is critical. However, AIV, which dones not have a D binding amino acid, still binds (not as well as the others, but it still binds enough to be selected), so we can see selectable binding can come about more simply, and via different pathways.
It can be even simpler than that. The hormone CGP binds to completely different amino acid’s in the ligand binding cleft, not DNKH at all, so there more than one way to get to bind specifically to a something as highly selective as a hormone receptor binding site. In terms of the lock and key mechanism, your claim, Dr. Behe, is that all tines of the “key” must be in place simultaneously to fit in the lock. But as we have seen above, this is not correct, even a very simple “key” will fit into the lock (keeping in mind both the lock and key are “floppy”).
All we need to get Darwinian selection of binding sites is to have weak but selectable binding. This can be accomplished with very simple changes, and a variety of unrelated structures, even for very highly restricted structures. Note again, while this example is small peptides binding to a receptor, a small peptide is in principle the same as a surface loop of a protein binging to another protein. Your model of highly restricted sequences which have to be in position simultaneously is just wrong.
Ian also thanks Behe for admitting that he was wrong and how he should apologize for how he treated ERV
I am pleased that you have acknowledged Vpu viroporin represents a real, de novo binding site.
Now if you had engaged with this in your response to Ms Smith, my respect for you would have risen immeasurably. To those of you not familiar with graduate and post graduate education, we actually want graduate students to disagree with us, robustly. After all, they are the ones carrying the torch of critical enquiry when we are gone. We don’t want them to accept our say so, “just because”. As scientists and educators our brief goes beyond just those PhD students we supervise, but to all engaged in critical enquiry, regardless of how we feel about their actual mode of delivery [1].
By “playing the man”[2] Dr. Behe, rather than engaging with Ms Smith’s arguments, you abrogated our responsibility as mentors and educators. Imagine the difference if you had dealt with Vpu Viroporin straight up. How about apologizing to Ms Smith now?
So Behe was given the opportunity to not only debate his side of the story but also apologize to ERV (Abigail Smith). What happened?
Infidel Guy, decided to correct this oversight and invited both ERV and Behe to present their position on his show.
Unlike the Christian show that let Behe dog me while they didnt even attempt to get me on their show, IG did not want his show to be me smacking on Behe behind his back, so they sent a second email.
Hi Dr. Behe- I apologize for sending a second request but I wanted to give you the opportunity to respond. Abbie Smith has agreed to be our guest on show to discuss ID and the arguments you have made for it. I believe you have had an ongoing dialogue with her in the cyber world and we wanted to get the two of you together to discuss these issues in real time. Abbie is scheduled to be on Thursday, February 7th at 8pm EST. However, if you wanted to be on we could re-schedule for a date that is convenient for both of you. Let us know of your interest and we will schedule you accordingly. Regards,
The response:
Hi, ___, sorry for not replying earlier. I appreciate your asking me, but I’m not interested in doing the show. Best wishes.
Mike Behe
‘Not interested.’
Well, I kinda figured you were ‘not interested’ in HIV research, LiLo, cause you didnt even Google ‘HIV’ before you decided to write a whole book on what it can and cannot do. Kinda figured you were ‘not interested’ in HIV research because your response to my essays was that the hard work of hundreds of scientists was ‘pathetic’. OH LOOK! More PATHETIC RESEARCH on Vpu in that PATHETIC journal, NATURE!
Yeah. Its the *research* thats pathetic, Behe.
Remind me to never anger ERV, she is a mighty force of science.
And that’s another day in the life of a DI fellow. Focus on Christian Apologetics and avoid any venues in which you will be confronted by real scientists. Is it not ironic that when ID talks about teaching the so-called controversy, it somehow seems to exclude itself? Now how dogmatic is that?
A final note is the following comment by Behe to Mark Chu-Carroll
Carroll cites several instances where multiple changes do accumulate gradually in proteins. (So do I. I discuss gradual evolution of antifreeze resistance, resistance to some insecticides by “tiny, incremental steps — amino acid by amino acid — leading from one biological level to another”, hemoglobin C-Harlem, and other examples, in order to make the critically important distinction between beneficial intermediate mutations and detrimental intermediate ones.) But, as Carroll might say, it is a non sequitur to leap to the conclusion that all biological features therefore can gradually accumulate. Incredibly, he ignores the book’s centerpiece example of chloroquine resistance, where beneficial changes do not accumulate gradually.
However this seems to be at odds with what the scientific papers do show.
For instance
Hastings, Bray and Ward (2002), writing in the journal Science, go into a little more detail. They note that sequential accumulation of mutations is the best explanation for chloroquine resistance:
The first mutations spread because they confer increased tolerance to CQ on parasites, enabling them to infect humans sooner after drug treatment — for example, mutation 4 allows parasites to infect people 6 days after treatment rather than 7 days. The relatively rapid elimination of CQ means that these are rather weak selective forces (6) and that the spread of these first mutations will be slow. Eventually, mutation 8 arises, which allows the parasite to survive therapeutic levels of CQ. Once above this threshold, the selective advantage conferred by this mutation becomes enormous and the pfcrt haplotype (now containing several sequentially acquired mutations) spreads rapidly across geographic regions where CQ is in common use. This appears to have occurred four times for CQ resistance: twice in South America, once in southeast Asia, and once in Papua New Guinea (see the viewpoint by Wellems on page 124) (10).
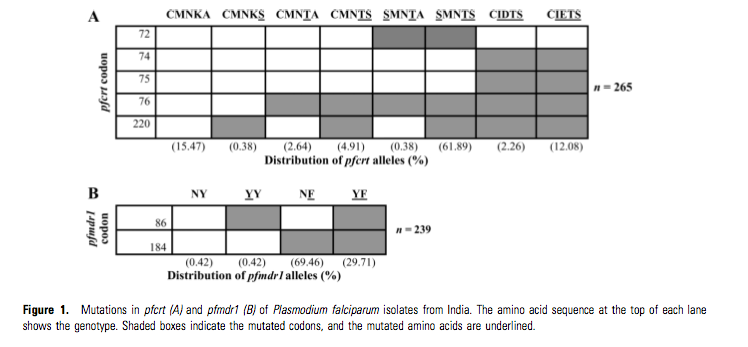
But this is hardly the only relevant paper, in “Progressive Increase in Point Mutations Associated with Chloroquine Resistance in Plasmodium falciparum Isolates from India”, Pooja Mittra et al conclude
Eight of our isolates also contained intermediate mutatedgene forms (7 isolates with the CMNTA genotype and 1 isolate with the SMNTA genotype), wherein a mutation was found at codon 76, whereas codon 220 had the wt allele (figure 1A). A study has shown that isolates with this intermediate form (the CMNTA genotype) show slightly higher IC50 values for CQ than do isolates with the wt allele [16]. Therefore, it has been proposed that acquisition of CQR is a stepwise process, and that association with additional mutations would give rise to a higher level of CQR.
 How long will it take for Behe to claim that what he said in his book, he did not really mean to say and that critics are once again misreading his book?
How long will it take for Behe to claim that what he said in his book, he did not really mean to say and that critics are once again misreading his book?
Blake Stacey’s collection of links (corrected)