
Meyer's Hopeless Monster, Part II
Note: I am extremely busy this summer, finishing grad school and moving to a postdoc. But when I got this book, I realized I wouldn’t be able to focus on my real work without having gotten my 2 cents in. This is a rough-and-ready piece, so typos and missing references, and missing explanations of technical terms are to be expected, although I’m sure they can all be figured out with a wee bit of googling. I am off to Evolution 2013 tomorrow and will be incognito, writing, after that. So I may not comment much. However I expect commenters to be reasonable discussants and polite and will ban people who break the spirit of this expectation. Cheers, Nick
Review of Stephen C. Meyer’s Darwin’s Doubt: The Explosive Origin of Animal Life and the Case for Intelligent Design
This week, a new book came out by Stephen Meyer, Darwin’s Doubt: The Explosive Origin of Animal Life and the Case for Intelligent Design. Having followed the ID movement and specifically its arguments on the Cambrian ‘Explosion’ for a long time, as well being somewhat up on the recent literature, and especially on phylogenetics, I feel that I have a pretty good sense of what to look for in any work purporting to be a capable commentary on the topic. As I read through Meyer’s book, though, in case after case I see misunderstandings, superficial treatment of key issues which are devastating to his thesis once understood, and complete or near-complete omission of information that any non-expert reader would need to have to make an accurate assessment of Meyer’s arguments.
After months of exuberant propaganda about the book from the Discovery Institute, I was half-expecting some kind of definitive, detailed, “serious” treatment of the issue from Meyer, who has had 4 years since his last book to work on this one, and has had 9 years since we raked over the coals his 2004 incompetent and self-plagiarized offering in the blogpost “Meyer’s Hopeless Monster”. I was sorely disappointed. I’ll hit a few of the main points. It would be nice if I had the time to write a comprehensive review, explain the issues from scratch in a Phylogenetics 101 sort of way, and provide detailed references, but given what my summer looks like, this is not likely. So, I’ll just outline what occurs to me as the most significant points– and what would occur to anyone else actually trained in phylogenetic methods, who also knows something about the Cambrian Explosion. If technical terms and the like don’t make sense, I encourage readers to google them. In the cases I have checked, Wikipedia does better at explaining the actual issues and methods than Meyer does.
.
A. THE “EXPLOSION” TOOK AT LEAST 30 MILLION YEARS, AND WAS NOT REALLY “INSTANTANEOUS” NOR PARTICULARLY “SUDDEN”
Darwin’s Doubt is festooned with illustrations, mostly redrawn from other sources in a rather strange cartoon-like format also found in other recent ID books. However, there is never an illustration like these:
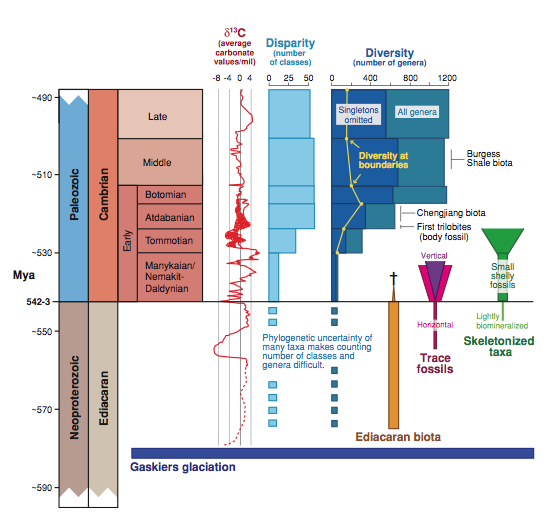 (Marshall 2006)
(Marshall 2006)
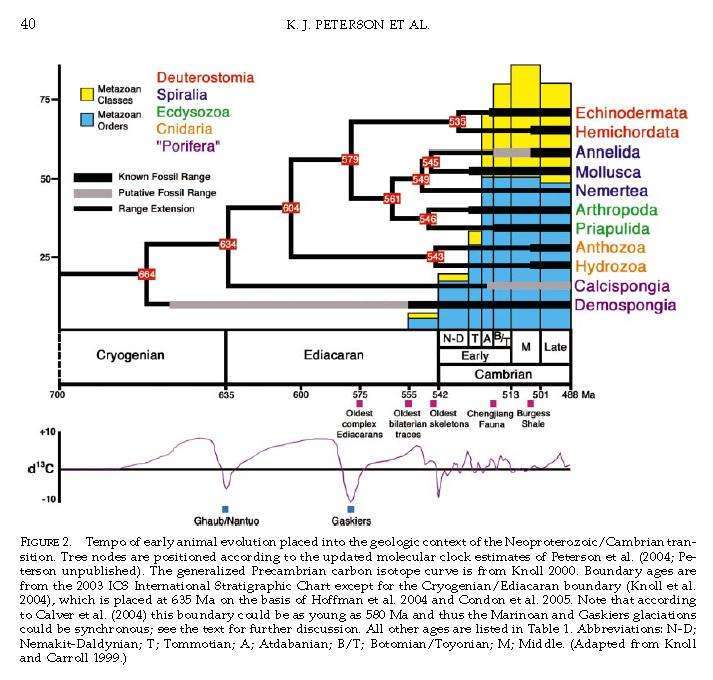 (Peterson et al. 2004)
(Peterson et al. 2004)
(Note: Erwin & Valentine 2013 of course contains similar illustrations)
Instead, we are treated to ultrasimple figures of the times of origin of “phyla”, which date back at least to the 1970s, although they’ve been endlessly copied by creationists/ID proponents and remain current in those circles because they convey the impression of “sudden” origin. Figures resembling this:
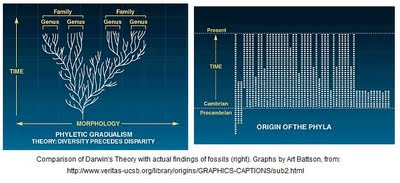
(source: originally from UCSB ID guy Art Battson, but he got it from earlier sources, and it has been copied in random places on the ID websites)
The reality is that, even on the most conservative interpretation of the fossil record which relegates all of the classic Ediacaran fossils to the stem below the bilaterian common ancestor, or to cnidarians, or to even more remote positions, we still have this sequence observed in the fossil record:
- 1. Before 700 mya, maybe well before: Single-celled eukaryotes (acritarchs)
- 2. Earlier Ediacaran: Multicellular animal eukaryotes, but simple, sponge-grade organisms
- 3. Later Ediacaran: Multicellular animal eukaryotes with more complexity, i.e. cnidarian-grade organisms
- 4. Very late Ediacaran: Simple slug-grade/worm-grade organisms (at least their tracks and burrows) – the first ones only making surface tracks and lacking burrowing ability. Making tracks suggests that the organisms have at least a front end and a back end, a mouth, anus, and gut connecting them. These are almost certainly bilaterians.
- 5. Very late Ediacaran: The very first biomineralized “skeletons”, e.g. Cloudina, basically a worm secreting a tube, as well as the first evidence of predatory boring. Cloudina gets no mention at all in Meyer’s book.
- 6. At the beginning of the Cambrian, we start to see more complex burrowing – e.g., vertical burrowing through sediment, clearly indicating worm-grade organization and an internal fluid skeleton, i.e. a coelom. The burrows gradually increase in complexity over 10 my.
- 7. Small shelly fauna:The shells, which started very small and very simple, gradually diversify and get more complex, radiating especially in the Tommotian. By the end of the Tommotion, some of the “small shellies” can be identified as parts of larger, “classic” Cambrian animals. The Tommotian is an utterly key period for any serious discussion of the Cambrian Explosion. Unfortunately, the word “Tommotian”, or any equivalent terminology (the detailed stratigraphy of the Cambrian is still being worked out, see Erwin & Valentine 2013 for a review), does not even appear in the book! The Small Shelly Fauna (SSF) gets just one (one!) mention in the book, buried in endnote 27 of Chapter 4, a whole chapter devoted to debunking the idea that the Ediacaran fauna is “ancestral” to bilaterians. (See discussion of the concept of “ancestral” below, which Meyer makes a complete hash of; however, I would tend to agree that the evidence is not good that the classic Ediacarans are within the bilaterian crown, as much because of the late date of #4-6, above, as anything.)
- 8. The earliest identifiable representatives of Cambrian “phyla” don’t occur until millions of years after the small shelly fauna have been diversifying, and they tend to be taxa on the stem below the crown of living phyla, rather than placeable within the crown. Trilobites are an exception, but what is often missed is that deposits like the Chenjiang have dozens and dozens of trilobite-like and arthropod-like organisms that fall cladistically outside of these respective clades. These are transitional forms! How can this fact not be highlighted!?!
- 9. In general, the earliest Cambrian relatives of the living phyla tend to be a lot more wormlike or sluglike than most modern representatives of the living phyla. Of course, many of the living phyla are basically still worms, and the more complex living phyla (e.g. molluscs, chordates) have early-diverging representatives or relatives that are rather more wormlike than the better-known representatives with more complex bodyplans. Even the earliest “fish” – actually either stem-group craniates, stem-group cephalochordates, or stem-group chordates – are basically filter-feeding worms that happen to swim. They don’t have jaws, scales, limbs, a bone skeleton, or anything else that most readers would associate with the word “fish”.
All of this is pretty good evidence for the basic idea that the Cambrian “Explosion” is really the radiation of simple bilaterian worms into more complex worms, and that this took something like 30 million years just to get to the most primitive forms that are clearly related to one or another living crown “phyla”, and occurred in many stages, instead of all at once. But, the reader gets very little of the actual big picture from Meyer.
.
.
B. MEYER DOESN’T UNDERSTAND PHYLOGENETICS, NOR MODERN PHYLOGENETIC TAXONOMY, NOR THEIR RELATIONSHIP TO OLDER TAXONOMY OF THE CAMBRIAN “PHYLA”
Much of Meyer’s discussion is framed by the Burgess Shale, the famous Middle Cambrian locality in Canada which yielded many well-preserved fossils. The interpretation and classification of the fossils found therein was a major topic throughout the 20th century. The history can (very!) roughly be summarized with the following:
- Stage #1: Charles Walcott (1850-1927) – Discovered the Burgess in 1909. Tended to lump the fossils in with the various modern phyla – e.g., anything that looks vaguely like an arthropod, call it an arthropod. Linnaean taxonomy reigns up until the 1970s.
- Stage #2: 1970s-1980s – In this period, we often see discussions of the Cambrian animals which are dominated by a weird kind of misbegotten mutant offspring caused by the attempt to fuse Linnaean ranked taxonomy with Hennigian thinking. In Linnaean taxonomy, taxa are defined basically by overall similarity, often relying on a few characters that are subjectively deemed “significant”. But with the onset of Hennig and thoroughly phylogenetic methods, the “phyla” start to be redefined to be monophyletic – thus removing, by definition, the whole idea of “ancestral phyla” – but initially, some people attempt to retain the meaningfulness of taxonomic ranks anyway. This has bizarre consequences, for example, if a cladistic analysis places a fossil even one morphological step outside of a “phylum”, then, by the logic of cladistics, it is a sister group to a phylum, and by the logic of ranked taxonomy, a sister group of a phylum ought to itself be a phylum. One can avoid this by expanding a “phylum” arbitrarily – thus moving you back to something like #1, above, and also camouflaging all possible fossils with transitional morphology by saying they are already members of existing phyla. Or you can create some paraphyletic, grade-based phylum which is ancestral to one or more modern phyla; but this violates the rule that taxa should be monophyletic, and also has the problem of arbitrariness, for where do you draw the line between one phylum and the next? Or, finally, you can start calling every stem group a new phylum, resulting in a profligation of “phyla” and “body plans” and the appearance of something really wildly different happening in the Cambrian than elsewhere. The epitome of the latter approach was Stephen Jay Gould’s (1989) Wonderful Life – crudely speaking, for Gould, if a fossil can’t be placed in a modern phylum, call it a new phylum, a whole new bodyplan, and build a whole theory of the contingency of evolution and our existence based on the resulting conceptual chaos of “fundamentally different” forms allegedly represented by the phyla.
(Another product of stage #1-stage #2-type thinking is the whole confused idea that in evolution, first we should see species-level divergence, then genus-level divergence, then family, etc., all the way up to phylum-level divergence. Meyer is clever in finding quotes from people, like Dawkins, endorsing this expectation, although all it shows is that they are products of being trained in stage #1 or stage #2, rather than in rigorous phylogenetic thinking, wherein it is recognized that ranks are retrospectively applied from the present, looking backwards.) - 3. 1980s-present – Gould’s popularization relied on the work of people like Simon Conway Morris and Derek Briggs, who were some of the early workers to apply Hennigian methods to Cambrian animals. Such methods are what show that many of the Cambrian animals do not fit within the living phyla, but rather they branch off of the stems below the crown groups. However, Gould was trained in the 1960s and never really “got” Hennig and cladistic methods in a serious way. But Briggs, Conway Morris, et al. did, along with hundreds of other scientists in other specialities that adopted cladistic methods and applied them to their domains of biology in the 1980s and 1990s.
All of the modern work on “transitional fossils” and the evolutionary origin of major groups (tetrapods, mammals, birds, whales, hominins, angiosperms, etc., etc.) is based on cladistic methodology and its successors (probability model-based phylogenetic methods). The study of the origin of the Cambrian “phyla” is no different. The major results of these studies, in taxon after taxon, with birds, mammals, tetrapods, hominins, etc. – as well as in well-preserved Cambrian groups – are that:
- 3a. The “key features” of the “crown groups” – groups defined as the clade containing two or more currently-living species, their common ancestor, and all of its descendants – did not come about all at once. Instead, when fossils are analyzed cladistically, we typically discover a bunch of species that morphological characters place below the crown – i.e., “stem groups”. The key thing about stem groups is that they have some, but not all, of the characters that make up the “body plan” of the crown clade under discussion. Among other things, this means:
- 3b. We can learn the basics of how “body plans” originated by using cladistics (or more sophisticated methods) to estimate the order and timing of each character change found in the crown group.
- 3c. The “body plans” did not originate instantaneously, all at once, unlike the ignorant contentions of creationists like Meyer.
- 3d. The “body plan” concept is basically just as arbitrary and flexible as the “phylum” concept – indeed, they basically both amount to a list of characters found in some monophyletic group (or, more commonly, a majority of the members of some phylogenetic group, as in any large phylogenetic group, some lineages get more complex while others get simpler, losing characteristic found in their ancestor – i.e., snakes are tetrapods despite having no legs). There is little or nothing to distinguish “highly significant” or “phylum-level” or “bodyplan-level” characters from other characters. For more on this, see my “Down with Phyla!” posts.
- 3e. The stem group taxa are the transitional forms that Meyer et al. are, allegedly, looking for.
- 3f. It is the step-by-step reconstruction of character changes that is the fundamentally important result that tells us about evolutionary history. This is the result that is closest to the data. The naming conventions are not of fundamental importance by comparison, even though creationists (including Meyer) usually distract themselves by focusing on names and taxonomic ranks rather than the distribution of characters.
- 3g. Once you’ve got 3a-3f worked out, then you are in a position to begin to talk in a sensible fashion about the evo-devo involved in causing particular character-state changes. Meyer is very good at misdirection – for instance, he will criticize cladistic methods for not answering the question of how new genetic information for character state changes comes about, and he repeatedly sets up bodyplan evolution as if it is evo-devo’s job to explain how each “phylum” got its “bodyplan” from scratch. But really, these approaches answer different, but complementary, questions. Cladistics breaks up the bodyplan characters and shows the basic steps they evolved in, and also which parts of the “bodyplan” are actually shared with other phyla. Evodevo then can work on each step, a much more manageable task.
Creationist “thinking” about the Cambrian, and “ID” thinking which derives directly and lineally from the 1980s creationist thinking on the topic, got all of its talking points (and figures!) from Stage #2 in the 1980s, and has continued basically unchanged since. No creationist/IDist has ever advanced to even understanding stage #3, let alone actually grappling with the implications, let alone even attempting to do the necessary work that would constitute a serious attempt at a rebuttal. Part of the problem is the residual influence of the confusing nature of stage 2 – many of the most prominent popularizers of evolution in the last generation, e.g. Dawkins and Gould, were educated in pre-phylogenetic times and do not always thoroughly grasp the implications. The other part of the problem is that, if creationists ever really did “get” stage 3, they’d have to give up the vast majority of the talking points and quote-mines they’ve been relying on for decades in their discussions of the fossil record, in the Cambrian and elsewhere.
A failure to grasp stage 3 is why Kevin Padian’s testimony at the Kitzmiller v. Dover case was so devastating on the topic of fossils, and why creationists/IDists will always fail, and fail miserably, when actually confronted with informed opposition in a format that allows detailed, extended discussion of the science. Unfortunately, a corollary of going to graduate school and gaining expertise in technical topics is that one becomes (a) busier and busier and (b) more and more cognizant that time spent rebutting creationists, who usually won’t put in the work required to understand you anyway, is time that should probably be better spent doing actual science.
Now, Meyer couldn’t stumble around in the Cambrian and phylogenetic literature without encountering a few of these issues. He does briefly discuss “lumpers versus splitters” and even Linnaean taxonomy versus rank-free taxonomy. But Meyer never presents for his readers the point that cladistic analyses reveal the order in which the characters found in living groups were acquired, nor the fact that stem taxa are the transitional fossils the creationists are allegedly looking for. And he especially avoids giving his readers any real sense of the number of transitional forms we know about for some groups, and the detail known about their relationships and about the order in which the characters of modern groups originated. The most egregious example is with the Cambrian arthropods and arthropod relatives. Why does Meyer never show his readers anything like this?
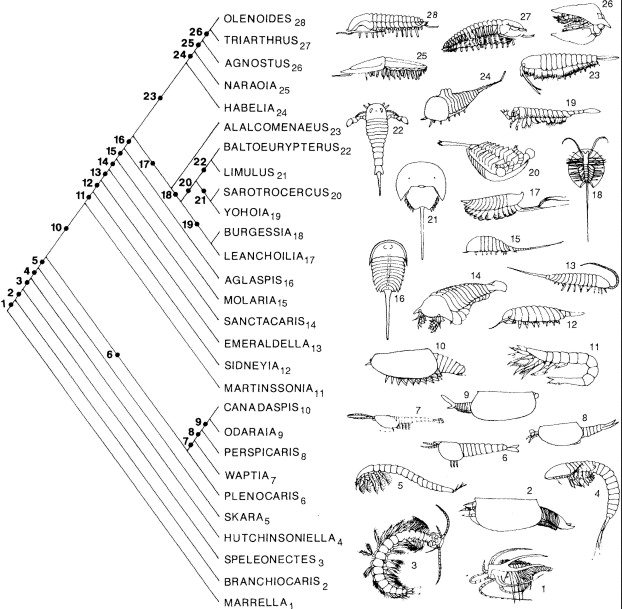 (source: Brysse 2008, originally Briggs & Fortey 1989)
(source: Brysse 2008, originally Briggs & Fortey 1989)
Or this?
.
C. NOT GETTING PHYLOGENETIC THINKING LEADS TO ALL KINDS OF OTHER PROBLEMS
Once you understand (B) and know something about the application of phylogenetic methods and thinking to the Cambrian animals, you will start to identify all kinds of other problems in Meyer’s book which stem from these fundamental misunderstandings. Again, the arthropods are instructive. Meyer continually and blithely refers to organisms such as Anomalocaris as “arthropods”, as if this were an obvious and uncontroversial thing to say. But in fact, anyone actually mildly familiar with modern cladistic work on arthropods and their relatives would realize that Anomalocaris falls many branches and many character steps below the arthropod crown group (see the figure above). Anomalocaris lacks many of the features found in arthropods living today. It is one of many fossils with transitional morphology between the crown-group arthropod phylum, and the next closest living crown group, Onychophora (velvet worms).
A related problem is Meyer’s treatment (mostly non-treatment) of “Lobopodia”, which he treats as a distinct phylum and includes in his phylum count. Meyer never spends a word on an actual critical discussion of what “Lobopodia” is supposed to mean – the term appears in a few picture captions, in the titles of some of his references, and in a quote of Simon Conway Morris. Whatever the method of naming the various scientists who use the term “Lobopodia” – Linnaean ranks, rank-free, etc. – as far as I know every authority would agree that lobopods are a paraphyletic grab-bag on the stems of the crown-group phyla Arthropoda and Onychophora (and perhaps also on the stem below their common ancestor). In other words, the arthropod and velvet-worm phyla evolved from lobopods, and lobopods contain a whole series of transitional forms showing the basics of how this happened! How anyone could write a book on the origin of Cambrian animals, without mentioning Cambrian Explosion 101 findings like this, is mystifying.
(Amongst other interesting implications of the above is the fact that the paraphyly of lobopods means that some of Stephen Meyer’s “phyla”, listed in endnote #5 of chapter 2, are actually subgroups of other “phyla” in the list! That, or Meyer accepts that one phylum evolved into two or more other phyla (Tardigrades may also be derived from lobopods; they are close to Arthrpoda and Onychophora on everyone’s analyses), which seems like an unlikely representation of his point of view.)
The above sorts of huge mistakes would not have happened if Meyer had taken care to get clear on the key concepts of “crown group” and “stem group”, which are absolutely necessary in any modern discussion of the Cambrian. Meyer does discuss briefly the crown-group versus stem-group issue, buried in endnote 5 of chapter 3, which discusses Budd and Jensen (2000). But Meyer’s discussion gets the definition of crown-group wrong, saying “crown groups arise whenever new characters are added to simpler, more ancestral stem groups.” No, crown groups are retrospectively defined by humans living in the present. We take two living taxa – say, humans and the platypus, and then whatever falls within the clade defined by these taxa and their common ancestor constitutes the crown group. The living “phyla” are just large crown groups – whether we call e.g. chordates a phylum or a superphylum or a subphylum, or dispense with ranks entirely, is basically a matter of taste. What matters is that this is a monophyletic group about which we can devise rigorous, testable hypotheses, such as the time of origin of this group, which fossils fall inside the crown or down on the stem, etc. We can also ask what fossils (and relationships with other living crown groups) tell us about the steps by which the character suite reconstructed for the last common ancestor arose, and how it later changed within the lineages within the crown.
Yet another confusion that Meyer exhibits relates to the idea of “ancestor”. As with all creationists, Meyer exhibits no understanding of the fact that phylogenetic methods as they exist now can only rigorously detect sister-group relationships, not direct ancestry, and, crucially, that this is neither a significant flaw, nor any sort of challenge to common ancestry, nor any sort of evidence against evolution. Distinguishing between a close sister-group relationship and an exact ancestor is just a level of precision that we cannot expect in most cases. It’s just a by-product of the method and the data available. (This is not quite the end of the discussion on this topic – eventually, we will have Bayesian methods that will assign probabilities to hypotheses of direct ancestry, although this will require formal definition and then data-informed estimation of what “ancestral lineage” means in terms of morphological variability within a lineage, the biogeographic and stratigraphic range of “morphospecies” through times, etc. End nerdy sidetrack.) But phylogenetic methods can and do regularly and rigorously identify collateral ancestry – sister group relationships, and ancestral grades and clades. We can say that birds descend from dinosaurs with essentially 100% statistical confidence, without knowing which if any currently-described fossils are exact direct ancestors rather than closely-related sister groups.
For all of the above reasons, almost every page of Meyer’s discussion of Cambrian organisms contains howlers of the first order. For example, in chapter 2:
First, the great profusion of completely novel forms of life in the Burgess assemblage (feature 3) demanded that even more transitional forms than had previously been thought missing. Each new and exotic Cambrian creature – the anomalocarids (see Fig. 2.10), Marrella, Opabinia, and the bizarre and appropriately named Hallucigenia – for which there were again no obvious ancestral forms in the lower strata, required its own series of transitional ancestors. But where were they?
Figure 2.10 is a picture of Anomalocaris.
Again, it is only by refusing to depict and specifically discuss of the inter-relationships of these sorts of taxa, and the data that supports them, and to mention the statistical support for the resulting relationships, that Meyer manages to pretend to his readers that these questions are not even partially answered, are unanswerable, and that “poof, God did it” is a better explanation. Here’s the cladogram from Legg et al. (2012) again:
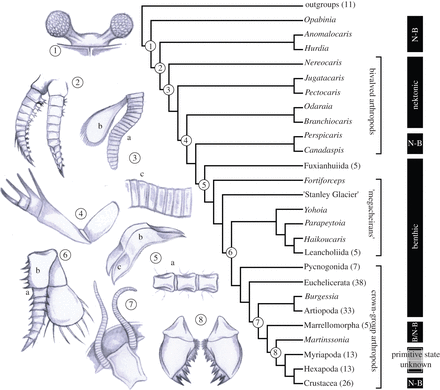
What goes into diagrams like this? They represent summaries of the morphological character data, which in this case you can see right here. Many readers, and virtually all creationists/IDists, will have little idea of the scale of effort that goes into constructing a dataset like this. These researchers, and the previous researchers that they are building upon, identified 580 individual, variable characters, each of which has to be identified, defined, divided up into discrete character states, and encoded. This laborious process had to be repeated for (in this case) 173 fossil taxa (correction – some are living, e.g. Drosophila). A lot of fossils are missing a lot of characters – typical and expected in paleontological analyses – but it is still a lot of work. After this, one runs a cladistic or other phylogenetic analysis (whole textbooks and courses are devoted just to this step of the process, and articles devoted to testing the reliability of phylogenetic methods, and improving the methods, are continually being published) and calculates support statistics. The support statistics are important since they tell you whether or not your data have any phylogenetic tree structure. Usually this doesn’t get major emphasis in scientific publications, because almost any biological dataset typically has extremely statistically significant tree signal, and this is true whether or not it agrees precisely with other analyses, and whether or not all relationships of interest to the researcher are precisely resolved with high support.
To anyone familiar with this work, it is simply laughable and pretty much insulting to see Stephen Meyer proclaim throughout his book that fossils with transitional morphology don’t exist, that the Cambrian body plans look like they originated all-at-once in one big sudden step. These statements don’t respect scientific process, they don’t respect the peer reviewed literature, they don’t respect the intelligence and knowledge of people who actually do know what they are talking about, they don’t respect the hard work of all the scientists that went out in the field and found these fossils, and then spent countless hours preparing them, describing them, inspecting them in microscopic detail, coding them in a morphology database, and analyzing them, all with care and effort and detail never taken by any creationist/IDist writer in any effort of comparative biology. And most importantly, Meyer’s statements don’t respect the data. They don’t follow the evidence wherever it leads, mostly because Meyer is ignoring most of the evidence.
.
D. IT GETS EVEN WORSE WHEN MEYER STARTS DISCUSSING PHYLOGENETIC CONFLICT
Despite the above omissions, Meyer is aware on some level that phylogenetic results, both on the molecular data from living organisms, and the morphological data from living and fossil organisms, represent some kind of challenge to his thesis. He thus devotes Chapter 7 and several other sections to dismissing the validity of phylogenetic methods and results in toto. His main argument is basically that phylogenetic results sometimes conflict, therefore the whole thing is meaningless. This exhibits a jaw-dropping level of incompetence. It’s amateurish in the worst possible way, the opening-your-yapper-without-knowing-the-first-thing-about-what-you-are-discussing sort of amateurism. There is no point in wading through Meyer’s mass of incomprehending quote-mining and cherry picking in detail, but in no particular order, here are some of the things that any attempt at an argument against phylogenetics and the phylogenetic evidence for the common ancestry of animals would have to address to merit any kind of serious attention from professionals:
1. Phylogenetic methods – the inference of phylogenetic trees from character data, whether molecular or morphological – are rigorous and well-tested. The easiest sort of test is computational. We can construct a phylogenetic tree in the computer, simulate the process of substitution (character change or mutational change) under one of hundreds of models (or a weighted combination of models, or some new model we just thought of), read off the simulated data at the tips, and then hand off this data to a phylogenetic inference program. We can then run the inference program, giving it no knowledge of the true tree, and see how well the inference program does at estimating the tree (and the evolutionary substitution model parameters as well, if desired). We can vary the models and model parameters and see when the methods break and when they work. The general conclusion from this research, over decades, is that the methods work quite well under a variety of conditions, and the areas where they don’t work as well are also reasonably well-known, as are the causes of weaknesses in inference, and these can be identified in many situations when they occur in real-world datasets.
2. If that weren’t enough, phylogenetic methods have also been applied to biological situations where the phylogeny is known – e.g., to viruses grown in lab cultures, when the cultures are manually divided and divided again. The methods also work well in reconstructing the actual history when the actual history is known for sure by direct observation.
3. “Tree structure” is not something wantonly imposed upon character data – it is something that is there or not, in quantifiable amounts according to standard statistics. The allegation that phylogenetics amounts to no more than “tree-drawing programs”, an allegation made in various places by the Discovery Institute’s Anne Gauger (without any rebuttal from Meyer or other DI colleagues) is an outrageous, incompetent statement that could only be made by someone with no familiarity, let alone coursework, in phylogenetics. It is easy to calculate statistics such as CI and RI, and compare them to CI and RI statistics calculated based on data reshuffled under a null hypothesis where any possible phylogenetic signal has been obliterated. (Well, it’s easy if you’re a phylogeneticist.) In virtually any real case, one will see substantial phylogenetic signal, even if there is uncertainty in certain portions of the tree. Any claim that the biological data shows no reliable tree signal about the base of the animal tree – a claim which Meyer makes repeatedly – would have to be a statistical, quantifiable statement. But Meyer shows no evidence of even being aware of statistical treatment of these issues.
4. “Conflict between trees” is also not an all-or-nothing thing, unlike what Meyer ignorantly suggests. The difference between two phylogenetic trees (or between 100 trees, or whatever) can be measured by statistics such as Robinson-Foulds distances. And, again, we can take the observed distance between two trees and compare it to a distribution of distances based on a null hypothesis of only random similarity between the trees. If the null hypothesis is falsified, then you’ve got strong statistical evidence for quantifiable agreement between trees, whatever the disagreements might be.
4a. Similarly, there are small disagreements and there are big disagreements. In cases where branches are short, there are few mutations or character changes grouping two lineages. Another dataset could easily support slightly different relationships, precisely because the branches were short and thus there was little time over which to average out the stochasticity of mutation and other evolutionary events. This is a minor disagreement. Whichever topology is the true one, the branching points of those lineages were close together in time, and it makes little statistical difference in an objective measurement of tree differences. If one dataset grouped arthropods with nematodes, and another dataset grouped them with plants, then we’d have a phylogenetic conflict worth making hay out of.
But to creationists/IDists, all phylogenetic conflicts of any sort are considered equally, crashingly devastating. It’s rather a lot like when the young-earth creationists argued if estimates of the age of the Earth varied between 4.5 and 4.6 billion years ago, this 100-my disagreement was huge, and therefore we should instead think the Earth is 6,000 years old.
5. Also, as mentioned in #1, phylogeneticists are well-aware of situations that are likely to produce uncertainty and disagreement between phylogenetic trees. These include:
- 5a. Long molecular branches without many sister groups (google “long-branch attraction”)
- 5b. Phylogenies with a low number of taxa (this includes many phylogenomic studies, because there are many fewer whole genomes than individual genes sequenced). This is just problem 5a on steroids. In general, I would prefer a phylogenetic study with one or a few well-conserved genes sequenced from 400 taxa well-spaced across the Animalia, rather than a phylogenetic study with only 10 taxa but with 400 or 4000 genes sequenced. Many of the phylogenetic conflicts that Meyer cites come from the latter sort of study. (And, in addition to the problems caused by poor taxon sampling, when an everything-and-the-kitchen-sink approach is taken with the genetic data, you are adding the well-known problems caused by poorly conserved, rapidly-evolving, harder-to-align sequences.) But, Meyer fails to alert his readers to this fact. Searches of the literature on terms like “taxon sampling” will be productive here.
- 5c. Studies based on one or a few characters are also problematic. The distribution of any particular character might disagree with some particular phylogenetic hypothesis for any number of reasons, including simply the stochasticity of the evolutionary process. (Actually, we can quantify the phylogenetic conservation of even single characters, and test to see whether or not the distribution of the character is really random with respect to phylogeny – often, even characters with a substantial degree of homoplasy have some phylogenetic signal.) The power of phylogenetic inference comes from the combination of many independent characters, ideally each with high initial probability of homology, as judged by alignability (for molecular data) or relative position and topology (for morphology). Ditto for studies based on the idea that character X is a “special character” that can only be gained and never lost, never has homoplasy, etc.
- 5d. With morphological data, the issue of clades vs. grades is important to understand, as is the issue of plesiomorphy. For example, Meyer takes the paraphyly of the sponges as some sort of amazing counterexample to phylogenetic consistency. But, sponges have always been at the base of the animal tree on anyone’s analysis, and changing this from being one deep side branch to two deep side branches on the way to bilaterians is not actually an amazing change, nor any huge contradiction of the morphological data.
Meyer just cites cases of phylogenetic conflict with reckless abandon, especially cases of 5a-5c, without any evidence of having any idea that causes of conflict are often well-known and can be predicted ahead of time. It is true that some of the problem is that many biologists who are not phylogenetic specialists are also sometimes naive about problems 5a-5c, and make overly dramatic statements when they get a slightly different topology than previous studies. Biology is a huge area, it’s impossible to be an expert on everything, and phylogenetic programs are easy to run but it takes a lot more detailed study to understand the details of what is going on. But surely, if someone is claiming, like Meyer, that the whole enterprise of phylogenetics, the common ancestry of Animalia, and the common ancestry of bilaterians are all bogus and based on nothing rigorous, then the onus on him is to know and review the ins and outs of phylogenetics with substantial expertise. Instead we get the typical beginner-level mistakes.
The reality of the phylogenetic situation with the Cambrian bilaterians is that a great many issues in animal phylogeny are much clearer than they used to be just a decade or two ago. Several long-suspected relationships based on morphology or development have been confirmed – e.g. echinoderms & chordates, or arthropods, tardigrades, and velvet worms. Major “superphylum” groups like lophotrochozoa and ecdysozoa are robust and here to stay. The monophyly of many of the classic “phyla” was also confirmed. Many of the other cases of “conflict” deal with groups that are basically worms which never had a huge amount of morphological support for any particular placements, or groups that may be reduced from a more complex ancestor because of small size, parasitism, or both (e.g. nematodes and relatives). This latter case of conflict also includes many cases where it turned out that one “phylum” nested within another, which should be impossible if, as creationists/IDists think, “phyla” are supposed to be highly distinct body plans that never grade into each other or evolve into each other.
6. Phylogenetics is a statistical science of estimation and approximation, like any statistical science. Like any measurement of almost anything interesting in science, data and inferences will have some degree of uncertainty and error. This is not some problem with phylogenetics, this is a common feature of all science, known to anyone who does science. Creationists/IDists, though, are almost unique in their penchant for categorical, binary, and inflexible thinking. They seem unable to think statistically, to realize that agreement and disagreement between hypotheses and data come in degrees, rather than all-or-nothing, and that it is quite possible for a general pattern of phylogenetic agreement to be extremely strongly statistically supported, while at the same time it being true that uncertainty remains about any number of detailed issues.
7. All of the major statistical phylogenetic issues I’ve raised above were put forward with much more patience and detail by Doug Theobald in his “29+ Evidences for Macroevolution” FAQ at talk.origins. Meyer cites this once, near the beginning of his quote-mining tour about conflict between phylogenies, but then asserts that “In reality, however, the technical literature tells a different story.” This just ain’t so. Meyer doesn’t really understand the technical phylogenetic literature, he doesn’t understand the difference between large and small phylogenetic disagreements, he doesn’t know that the similarity between phylogenetic trees can be measured and that different datasets usually produce trees with a high degree of overall similarity, and he doesn’t understand the statistical methods that tell us that the raw data, whether molecular or morphological, typically have a high degree of tree structure. He doesn’t even get to addressing the basic material covered in Theobald’s FAQ, let alone the technical literature cited therein, which is specifically devoted to testing common ancestry and phylogenetic consistency, tests which are passed with flying colors in a statistically rigorous way in case after case. Meyer’s contribution to the phylogenetic portion of this discussion is therefore without merit and worthless, at least to anyone who knows something about phylogenetics.
E. OTHER ISSUES
Ironically, despite making such a hash of the key issues for dealing specially with the Cambrian fossil record and the phylogeny of animals, most of Meyer’s book is about other topics entirely. My guess is that Meyer sensed on some level that he actually was on weak ground basing his argument on the fossil record and phylogenetics, which in reality are the home turf of evolutionary biologists and both of which actually tell strongly against his argument to anyone who is actually familiar with the issues I have reviewed.
Instead, Meyer’s main argument is really about “information”. Back in his Signature in the Cell book, Meyer asserted that the only known source of information was intelligence, and that therefore we could safely infer that intelligent design was behind the origin of life. This is problematic for all sorts of reasons, but one of the biggest was that intelligence is not the only known source of information – in particular, evolutionary processes of mutation+selection can produce it, thus intelligence is not the only cause of information, thus “information” isn’t some magical signal uniquely evidencing intelligence in the complete absence of any other evidence. This objection was particularly devastating to Meyer’s argument in Signature because of the way Meyer based his argument on mere “information”, rather than primarily on the difficulty of explaining the origin of life or some such.
Meyer’s response to the but-evolution-produces-information counterargument was to argue that this didn’t answer how information came about during the origin of life, which happened before there was evolution. This actually isn’t necessarily strictly true – look up “prevolution” and Addy Pross – and, anyway, the vast majority of Meyer’s presentation of the information ==> intelligent design argument explicitly relies on the premise that information is uniquely and exclusively produced by intelligence. Furthermore, it was clear enough in Signature, and in the rest of Meyer’s writing, that he thinks, in a quite simple-minded way, that all genetic information everywhere in biology is produced by intelligence, and that evolutionary processes cannot do it at all. For Meyer, the origin of life, the origin of Cambrian groups, and the origin of each and every gene are all the product of one thing – divine intervention, barely disguised with the weasel words “intelligent design”. Thus, for several reasons, any and all evidence for the evolutionary origin of new information really was quite relevant to the assessment of Meyer’s argument in Signature.
But, whatever one thinks about Meyer’s argument in Signature about information and the origin of life, and his avoid-the-topic-of-evolution defensive strategy there, there was no way for Meyer to get around the but-evolution-naturally-produces-new-genetic-information argument in Darwin’s Doubt. Meyer is now in the realm of animal evolution, where there is absolutely no debate about whether or not evolutionary processes are a potential answer to the “where did new genetic information come from” question. Here, finally, Meyer has to confront the critics who have been pounding him on this issue ever since the 2004 PT post “Meyer’s Hopeless Monster”.
Longtime PT readers may remember that the “Meyer’s Hopeless Monster” post was a critical review of Meyer’s article on the Cambrian Explosion (well, sort of on the Cambrian Explosion) in The Proceedings of the Biological Society of Washington (D.C.). PBSW was a rather obscure journal devoted mostly to alpha taxonomy, but it was edited by Richard von Sternberg, who I believe is now on the Discovery Institute payroll. The year 2004 was just about the peak of the ID movement, and after we put up our critical review, stating that Meyer’s article was substantially self-plagiarized from his previous works, was wrong on a large number of specific issues, and thus could not be considered a competent work of biology, and the peer-review couldn’t be considered competent either, a rather impressive fracas started. The whole story can be reviewed in endless detail in the PT archives.
We never got a detailed response from Meyer, and a promised Discovery Institute detailed rebuttal flamed out after one or two posts. But now, at long last, Meyer devotes a big chunk of chapter 11 to rebutting one of the key arguments we made, namely that the origin of new genetic information by evolutionary processes is well-documented and well-understood, and this is demonstrated in part by a nice detailed review article by Long et al., (2003), Nature Reviews Genetics, entitled “The origin of new genes: glimpses from the young and old.” The paper has two nice big tables, one giving a review of the many mutational mechanisms (all of them mutational processes known to occur naturally and in the lab) involved in the production of new genes, and another giving examples reviewing papers that had reconstructed the origin of new genes, typically recent genes where the evidence was very good and very clear, although this wasn’t the case for every single example. The paper was so handy that it was used again as an evidential exhibit in the Kitzmiller v. Dover case, and it even got cited by the Judge in the final decision as shooting down the ID argument that genetic information requires intelligence.
All of this apparently stung Meyer and the Discovery Institute folks, although it took them a long time to muster any sort of response to this fatal counterargument. Five years later, in 2010, we finally got a response from Casey Luskin, which basically amounted to the juvenile assertion that invoking mutations (which are known, regularly observed natural processes, which leave easily detectable and obvious evidence in the genome) and natural selection (which is also a known, observed natural process, and which also often leaves detectable statistical evidence in the genome, although it is not always quite as easy to detect as mutations) amount to “waving a magic wand” and telling a vague “just-so story.” Please turn off your irony meters before you remember that this is coming from an ID advocate, who literally thinks that “poof, God did it through divine intervention!” should be an acceptable scientific explanation. Luskin’s other argument seemed to be that if we don’t know everything about the new gene in question – its exact function and the exact reasons that selection occurred – then we know nothing at all, and can safely sweep under the rug all of the evidence that we do have for the mutation and selection explanation (for example, in the case of Sdic, which I reviewed here with pictures). Why he expects this argument to be convincing to any serious scientist who works in genomics, genetics, or similar fields mystifies me. We know a lot about a lot of things in biology, but there is basically nothing about which we know everything we would like to know. Science progresses nonetheless, and we can say many things with confidence nonetheless, in part because we have rigorous statistical methods to detect past selection, to reconstruct the relationships of genes, and to reconstruct the approximate sequence of mutational events to some recently-originating new gene.
(Also note that Richard Hoppe pithily pointed out the scientific implications of Luskin’s “invoking well-known, well-understood natural processes is equivalent to just waving a magic wand and telling a just-so story” argument, in this PT post)
Given Luskin’s embarrassing attempt, does Meyer do any better in 2013 at rebutting Long et al. and similar copious evidence that new genes can and have been regularly produced by natural evolutionary mechanisms? Sad to say, nope, he doesn’t. Meyer first reviews his 2004 paper and the “Meyer’s Hopeless Monster” episode. (Inaccurately, as it turns out – Meyer seems to say that “Meyer’s Hopeless Monster” was first published on the “atheist” (?) talkreason.org website, and that it was published late in the controversy over Meyer’s PBSW article; in fact, if memory serves, the article was published here on PT first, and furthermore it was the very first public notice that anyone made of the Meyer article.) Meyer then just half-heartedly lists a couple of random complaints and a couple of vague and question-begging alternative explanations of the data. Namely:
- 1. Hypothetical reconstructions are not data (never mind that the ID claim is that no way, no how, can natural evolutionary processes produce new genetic information, and that information is uniquely produced by intelligence; and never mind that all of science is about proposing and testing hypotheses. Meyer’s tactic in this chapter of emphasizing words like “postulate” and “suggest” is just juvenile, no better than what you get from the young-earth creationists who just fell off the turnip truck, to repurpose a description of YECs that Meyer used to like to use).
- 2. The common ancestral gene before a gene duplication is inferred rather than observed (never mind phylogenetics, and all of the evidence for its reliability and rigor mentioned above; also, never mind the fact that in recently-evolved cases, i.e. in Drosophila, we sometimes have the exact or near-exact original ancestral gene sequence present; also, never mind that gene duplications have been observed in the lab, and natural polymorphisms in gene copy number are regularly observed in natural populations, including in humans; never mind that the mutational mechanisms producing gene duplications, e.g. unequal crossing over, are also well-understood and sometimes directly observed).
- 3. These papers assume the existence of an ancestral gene (in fact, the title of Meyer’s Chapter 11 is “Assume a Gene”). Never mind that, when it comes to the Cambrian bilaterians, we have oodles of evidence for prexisting animals (cnidarians and sponges, both of which have lots of genes shared with other animals) as well as single-celled eukaryotes, which have plenty of genes, further back. Never mind that Meyer’s information argument hangs completely on it being a universal generalization with no exceptions that intelligence is the only source of new genetic information. Never mind that in any process of tracing a gene’s history back in time the trail will eventually go cold – it is a necessary result of mutational decay of sequence similarity – and this is not evidence of any miraculous process at the beginning of it, just evidence that human knowledge is limited.
- 4. Meyer rather randomly suggests that maybe convergence is an alternate explanation of the similarity of genes normally interpreted as duplicates. This is laughable even in the most extreme cases of molecular convergence that Meyer can find (a hearing-related gene in bats and whales, which amount to maybe a dozen convergent sites out of hundreds, if memory serves).
- 5. Then Meyer drags in this whopper: “it is possible that similar genes might have been separately designed to meet similar functional needs in different organismal contexts.” (italics original). Really? Does Meyer really think that The Designer monkeyed with the genome of one Drosophila species among thousands to produce sdic, and just happened to place it in between the two genes that look like they provided the ancestral gene chunks, and just happened to litter the genome region with other telltale sequence bits that indicate a typical mutational process, and then did some targeted reduction in genetic diversity to make it look like a selective sweep recently happened? Really? This is no better that the Omphalos argument that God made the Earth appear to be billions of years old, but actually it’s only 6,000 years old.
- 6. As a final point, I would note that even Michael Behe accepts the evolutionary origin of mere new genes with different functions – even ones that are now essential to a species, and which cause death if deleted! See his June 2013 post, “A new gene fits in without a ripple”! Complete and utter abject defeat for the IDist evolution-can’t-produce-new-information argument! (And really, for IC as well, but that’s another topic.)
Perhaps dissatisfied with his phoning-it-in response to the work of hundreds of scientists documenting the origin of new genes (work which has, by the way, greatly expanded since 2003…even writing a review article now would be a much bigger task), Meyer then switches topics to ORFans. Here he repeats the usual IDist problems with complete ignorance of the relevant statistical issues involving ORFans and homology searches between genomes – as, unfortunately, can be found in some of the scientific literature on this topic.
Meyer’s last argument basically reverts to the improbability argument – sequence space is large, functional space is small. This trumps all of the above evidence, according to Meyer. It never occurs to Meyer that his assessment of the probability of functional sequences might just be wrong, and that the deluge of evidence that new genetic information is easy to evolve is pretty direct evidence that his probability assessment is wrong.
Elsewhere in the book Meyer invokes some other arguments to justify the “improbability of functional sequence” argument. These are Behe & Snoke’s 2004 argument about multiple simultaneous mutations, Behe’s Edge of Evolution argument about “chloroquine complexity clusters” and protein-protein binding sites, and work by Douglas Axe and Anne Gauger, run out of the Discovery Institute’s approximately 2 person research institute, the Biologic Institute, which typically involves making evolutionarily absurd modifications to proteins and then showing that they don’t work. This latter stuff is actually about as conceptually bad as the old YEC idea that evolution meant there should be such a thing as a “crocoduck”.
Anyway, most of this has been rebutted elsewhere on PT, and there is little point in doing it again. It is pretty strange, though, that most of these talking points were invoked in a very similar way in last year’s DI book on human origins – it looks like they will throw in some Behe and Axe just about anywhere. So really, when you buy a book from the Discovery Institute, you only get a portion of material devoted to the topic, and the rest is just a rehash of the same unconvincing material about what-if-multiple-mutations-were-required and poorly-informed intuitions about the improbability of evolutionary processes.
The multiple-required-mutations stuff, by the way, is basically just Behe’s refuted “irreducible complexity” argument disguised as an argument about sequence evolution, and is only relevant if it can be shown that 2 or more neutral mutations ever were required for anything relevant to the Cambrian Explosion, but, as is typical in DI literature, this is just blithely assumed rather than argued for. Showing it for any case would be nontrivial, and every detailed study I have read about multiple-mutation adaptations indicates that it doesn’t usually apply – instead, what typically occurs to produce an alleged “multiple mutations required” adaptation is that a variety of single-step mutations are selected as partial, imperfect adaptations to some chemical or environmental stressor. One or more of these enables the viability of some mutation of high adaptive value, which is then followed by selection of further mutations ameliorating whatever negative impacts the “major” adaptive mutation might have incurred. At the end of this process, you have something that would look as if several mutations would have had to happen at once, but only if you are ignorant of the somewhat circuitous-but-adaptive path (sometimes with selection for functions other than the final one) that was actually taken.
Meyer’s other go-to argument on information is basically, “even if evolution can produce new genes, it can’t produce new protein domains”. This is (A) basically a tacit admission of defeat on the information question, and (B) there’s no evidence that new protein domains were required in the Cambrian – I’d be surprised if any protein domains are known that are both unique to and required for the existence of Animalia. Animal genomes mostly just elaborate on and expand (through duplication) the already-complex pre-existing eukaryote genome. (For a tiny contribution to the where-eukaryotes-came-from question, you might check out my new coauthored PNAS paper, out this week.) The basic protein domains mostly originated very far back in evolutionary history, where things like ultrahigh population sizes help out, as well as the fact that there are probably a fairly limited number of protein domains, and the fact that point mutations and rearrangements can, on occasion, produce new or modified protein folds, in some cases without having much effect on enzymatic activity.
.
CONCLUSION
Meyer discusses a number of other issues, e.g. development, that would be better treated by others. And he spends a lot of time on the usual quote mines, the misrepresentation of evo-devo, self-organization, Punk Eek, the Altenberg 16, etc., all as “alternatives” to “Neo-Darwinism”, all of which is mistaken and misshapen but which would take longer to untangle than I have time for in the near future. He also repeats the usual ID talking points about junk DNA and the ENCODE project, apparently completely ignorant of the devastating responses based on the huge variability in animal genome sizes, amongst other issues.
Even without addressing all of these other issues in depth, I think the above shows that Meyer’s book is already holed beneath the waterline on the key issues of Cambrian paleontology, phylogenetics, and the information argument. I’m not sure it deserves much more of anyone’s time. Sadly, some vaguely respectable people seem to have ignored the crashingly obvious flaws and endorsed the book, although in at least some cases they are already known for promoting bizarre opinions in other contexts. Enthusiastic reviewers in the blogosphere, like Tom Gilson at Thinking Christian, seem to lack even Wikipedia-level research abilities in critically assessing Meyer’s claims.
The one refreshing bit of the book is at the end, where Meyer basically admits that, yes, this really is all about bringing an interventionist God back into science, and thereby reconciling and harmonizing science and religion, and solving the problems of meaning in the culture and belonging in the Universe, or something. How exactly this could ever work, even if Meyer’s argument’s succeeded, is not explained. Meyer completely and explicitly punts on the question of providing any sorts of answers on what exactly is supposed to have happened at the Cambrian or anywhere else in geological history, on the ID view. All we get is ID did something, somewhere, somehow, for some reason, never mind extinction, the millions of years of twiddling around with arthropods, the billions of years of twiddling around with bacteria, the endless examples of apparent evidence for evolution, etc. If Meyer takes his own arguments at all seriously, he is invoking divine intervention not just for the origin of life and the Cambrian, for basically every new gene, ORFan, any adaptation of any significance, and some ill-specified level of morphological difference. This is, probably, billions of separate divine interventions. It essentially amounts to invoking divine intervention at every instance where Meyer personally doesn’t understand something, even in cases where scientists understand something quite well, and Meyer simply can’t be bothered to do the work necessary to understand what they are talking about. As I’ve said before, the real problem with creationists/IDists isn’t when they stick God into the gaps in current scientific knowledge. Such a thing is unwise, given history, but at least questions that all of humanity still wonders about are vaguely worthy of divine intervention. The real problem is when creationists/IDists insert God into the gaps in their own personal knowledge, gaps which have already been filled by scientists. Here it is completely clear that the creationists/IDists are arrogant enough to call God down from Heaven to cover for their ignorance, basically because they are unwilling to do the basic “due diligence” and hard work required to get a basic understanding of the topic they commenting on. I’m not sure that most long-lived religious traditions actually support that kind of behavior.
A final thought: I’d like to see Meyer, or his defenders, explain which of the lobopods and stem arthropods and arthropods in the figures above are in different “genera”, “families”, etc., up to “phyla”, and why. I think that if you plunked those fossils down in front of an ID advocate without any prior knowledge except the general notion of taxonomic ranks, the ID advocate would place most of them in a single family of invertebrates, despite the fact that phylogenetic classification puts some of them inside the arthropod phylum and some of them outside of it.
.
.
References (originally provided here)
Briggs, D.E.G.; Fortey, R.A. (1989). “The early radiation and relationships of the major arthropod groups.” Science, 246(4927), 241-243. http://www.sciencemag.org/content/246/4927/241
Brysse, Keynyn (2008). “From weird wonders to stem lineages: the second reclassification of the Burgess Shale fauna.” Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences. 39(3), 298-313. http://dx.doi.org/10.1016/j.shpsc.2008.06.004 , http://www.sciencedirect.com/science/article/pii/S1369848608000393
Budd, G. E., and S. Jensen. 2000. “A critical reappraisal of the fossil record of the bilaterian phyla.” Biological Reviews 75:253-295.
Erwin D and Valentine J (2013). The Cambrian explosion : the construction of animal biodiversity. Greenwood Village, CO, Roberts and Company Publishers. Link: http://www.amazon.com/The-Cambrian-Explosion-Construction-Biodiversity/dp/1936221039
Legg, David A.; Sutton, Mark D.; Edgecombe, Gregory D.; Caron, Jean-Bernard (2012). “Cambrian bivalved arthropod reveals origin of arthrodization.” Proceedings of the Royal Society, Series B: Biological Sciences. 279(1748), 4699-4704. http://dx.doi.org/10.1098/rspb.2012.1958 http://rspb.royalsocietypublishing.org/content/279/1748/4699/F3.expansion.html
Marshall, Charles R. (2006). “Explaining the Cambrian ‘explosion’ of animals.” Annual Review of Earth and Planetary Sciences. 34: 355-384. http://dx.doi.org/10.1146/annurev.earth.33.031504.103001
Meyer SC (2013). Darwin’s doubt : the explosive origin of animal life and the case for intelligent design, HarperOne. Link: http://www.amazon.com/Darwins-Doubt-Explosive-Origin-Intelligent/dp/0062071475
Peterson, Kevin J.; McPeek, Mark A.; and Evans, David A. D. (2005). “Tempo and mode of early animal evolution: inferences from rocks, Hox, and molecular clocks.” Paleobiology 31(2Suppl), 36-55. http://dx.doi.org/10.1666/0094-8373(2005)031[0036:TAMOEA]2.0.CO;2 E.g. as cited by Kevin Padian in the _Kitzmiller case: http://www.sciohost.org/ncse/kvd/Padian/Padian_transcript.html#s031
.
.
Strongly recommended reading (and the references therein)
Alan Gishlick, Nick Matzke, and Wesley R. Elsberry (2004). “Meyer’s Hopeless Monster.” Panda’s Thumb post, August 24, 2004. http://pandasthumb.org/archives/2004/08/meyers-hopeless-1.html
The “Meyer 2004” Medley - The Panda’s Thumb – the complete history of the Meyer 2004 craziness.
Matzke, Nicholas (2005). Down with phyla! - The Panda’s Thumb, which reviewed:
- David Fitch and Walter Sudhaus, “One small step for worms, one giant leap for ‘Bauplan?’” Evolution & Development 4:4, 243-246.
- Budd, G. E. and S. E. Jensen. 2000. “A critical reappraisal of the fossil record of the bilaterian phyla. Biological Reviews of the Cambridge Philosophical Society 75:253-295.
- Graham Budd (2001). “Climbing life’s tree.” Nature 412, 487.
- Jaume Baguñà And Jordi Garcia-Fernà ndez (2003). “Evo-Devo: the Long and Winding Road.” Int. J. Dev. Biol. 47: 705-713. PubMed
- Walter Sudhaus (2004). “Radiation within the framework of evolutionary ecology.” Organisms, Diversity & Evolution 4, 127-134.
- Gonzalo Giribet (2003). “Molecules, development and fossils in the study of metazoan evolution; Articulata versus Ecdysozoa revisited.” 106: 303-326.
Matzke, Nicholas (2005). Down with phyla! (episode II) - The Panda’s Thumb
Matzke, Nicholas (2007). Meet Orthrozanclus (down with phyla!) - The Panda’s Thumb
(A late edit: I made a local copy of the Briggs & Fortey 1989 / Brysse 2008 figure, as it was hosted on a journal website and might not load for everyone depending on proxy settings etc.)